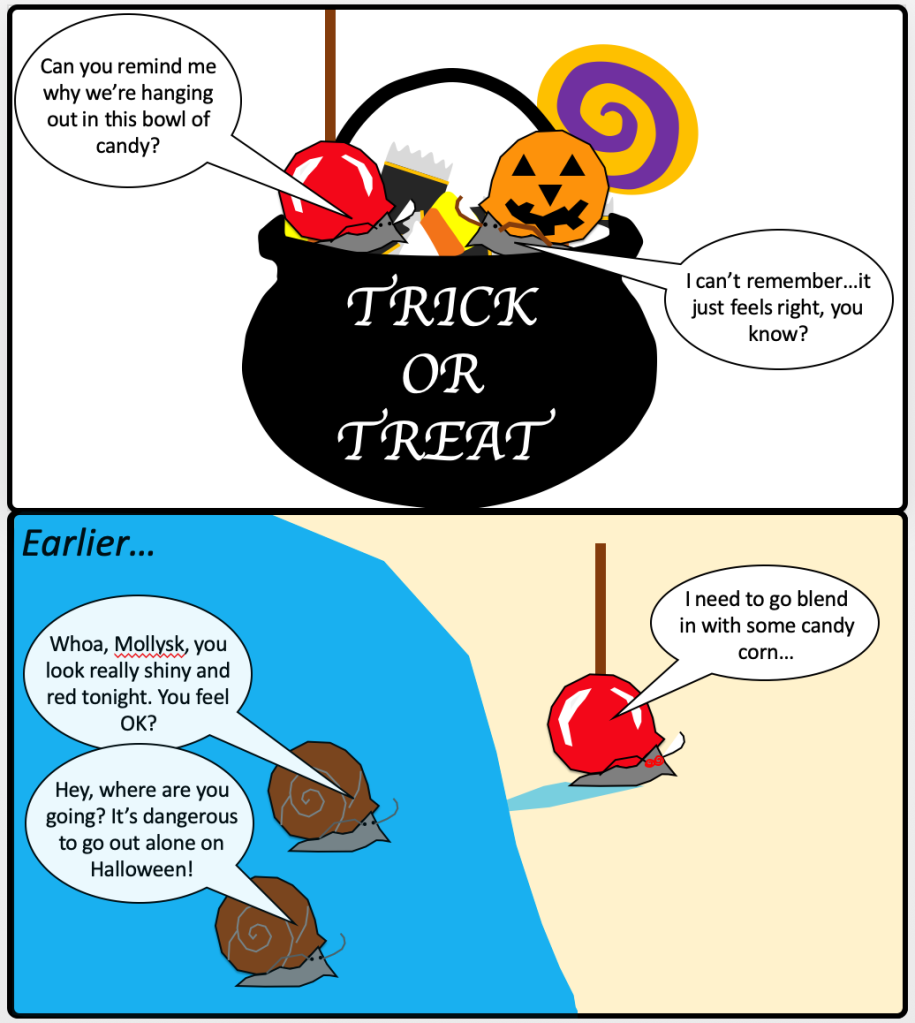
Happy Halloween! Turn on some spooky music, grab a handful of candy, and settle in for a spooky post about host manipulation by parasites! But do be careful eating; you never know when a parasite might be sneaking into your diet.
What is adaptive manipulation?
If you’re a parasite enthusiast, you’ve probably heard about parasites that manipulate host behavior. But what, exactly, is host manipulation? We expect that infected hosts will often act “sick”; they might try to fight off their infections by raising their body temperature (fever) or eating medicinal plants, or they might be suffering from so much pathology from their parasites or immune responses that they lose their appetites, act lethargic, etc. But none of those sickness behaviors are adaptive manipulation. To be adaptive for the parasite, the changes to the host’s phenotype (coloration, morphology, and/or behavior) must produce a fitness benefit for the parasite, usually by increasing parasite transmission from one host to the next. Following this definition, the host’s phenotype is actually the parasite’s extended phenotype: the parasite’s genes control how the host looks or behaves.
To show that a host’s phenotype is an example of adaptive manipulation, we need to show that that the phenotype actually increases parasite transmission. This can be tricky! But here’s an example. Last week, we talked about the life cycle of Euhaplorchis californiensis, a trematode with a complex life cycle. The trematode infects killifish as the second intermediate host; specifically, it infects the hosts’ BRAINS. Infected killifish are more likely to do flashy swimming behaviors, like dashing up to the surface. This seems like an adaptive manipulation that would make infected fish more likely to get eaten by their definitive bird hosts; but how would you prove that? If you’re Kevin Lafferty, you’d put a bunch of killifish in two cages in the salt marsh, leaving one open to birds and one closed to birds. You can hear Kevin describe the experiment in this TED talk. At the end of the experiment, the cage that was open to birds had fewer infected fish than the cage closed to birds, because the infected fish with the flashy behaviors were more likely to be eaten than uninfected fish. The manipulated behavior increased transmission by increasing predation.
Most parasites that manipulate host behavior are not as well studied as Euhaplorchis californiensis. Therefore, we should be careful not to overstate adaptive benefits where they do not exist or have not been demonstrated, or to overdramatize demonstrated adaptive behaviors. For example, we often say that crickets infected with nematomorphs commit suicide by jumping into water, but not all crickets die or even lose the ability to reproduce after being infected. Furthermore, infected crickets aren’t so water-fixated that they hop in a straight line to the nearest waterbody; they’re just more likely to end up in water than uninfected crickets. It’s important to remember that many details tend to be glossed over when telling exciting and spooky stories, including those about parasites that manipulate host behavior.
What kinds of manipulation exist?
We’ve mentioned a few examples, but how common is adaptive manipulation among parasites? We don’t know exactly how many parasites have evolved to manipulate their hosts, but we do know of a few hundred examples spread across many parasite groups (platyhelminths, acanthocephalans, nematodes, nematomorphs, arthropods, bacteria, fungi, protozoa, and even viruses). In fact, host manipulation mechanisms seem to have evolved independently many times within and between parasite groups. From this, we can conclude that adaptive manipulation is common, despite seeming so biologically bizarre.
But of course, there is spectacular variation among parasites in the ways that they manipulate their hosts. We can divide these methods into roughly five manipulation targets: predator avoidance, habitat choice, feeding or foraging, bodyguard behavior, and contact rates. I’ll explain each below.
Predator avoidance. Host manipulation seems especially common for parasites with complex life cycles that involve trophic transmission, where an intermediate host is consumed by a definitive host. In these systems, parasite transmission may be increased whenever the intermediate host becomes more conspicuous or less defended from the definitive host. Euhaplorchis californiensis is a good example of a parasite the manipulates how prey behave in the presence of predators.
Habitat choice. Sometimes parasites with complex life cycles have host species that do not share the same habitats or propagules (environmental stages) that require different habitats than the hosts, making it tricky for the parasite to make it from one host to the next. For example, larval nematomorphs infect terrestrial crickets and other arthropods, adults are free swimming in aquatic environments, and then larvae need to make it back to terrestrial environments to infect more arthropods. Nematomorphs manipulate cricket behavior to make them more likely to jump into water bodies—habitats that they would not usually choose. The story for how nematomorphs make it back to terrestrial environments might be even weirder, but the details are still being studied.
Host feeding behavior. Many parasites are consumed and/or transmitted during feeding, like those that are transmitted by blood-feeding insect vectors. If a vector only ever takes one blood meal, it cannot move parasites from one host to the next. To increase transmission opportunities, several parasites are known to affect vector feeding behavior, causing vectors to feed on more hosts and/or change their feeding behavior to increase the likelihood that a given feeding event will lead to transmission. A very different example might be trematodes that castrate their first intermediate snail hosts; snails reduce the time and energy that they spend reproducing when they are castrated, and instead spend that time eating, growing larger, and making more tissues available for the parasites to turn into more parasites. You can more read about parasitic castrators and gigantism in this blog post, including whether gigantism is truly adaptive for parasites.
Bodyguard behavior. Some parasites cause their hosts to co-opt normal behaviors or use abnormal behaviors to protect their parasites. For example, parasitoid wasp larvae cause orb weaving spiders to construct a protective web, and after eating the spider, the wasp larvae use the protective web as a safe hide out while they pupate. If you don’t go Google parasite bodyguards after reading this, you aren’t doing Halloween right.
Contact rates. Finally, although parasites with complex life cycles are usually the focus of adaptative manipulation research, parasites with direct life cycles and direct transmission might also benefit from manipulating host behavior. For example, if sexually transmitted parasites can cause their hosts to be more promiscuous, the parasites will be more likely to make it to a new host. It is unclear if there are few examples of this type of manipulation because it is understudied or because it is uncommon among directly transmitted parasites.
Now that you know what adaptive manipulation is and isn’t and what types of manipulation exist, you’re ready to go explore the many amazing examples that scientists have discovered or maybe to go discover a new example of your own! If you’re looking for good places to start, I recommend this TED talk by Ed Yong and this cartoon by The Oatmeal.
References:
Moore, J. Parasites and the Behavior of Animals. (Oxford University Press, 2002).
Poulin, R. Chapter 5 – Parasite Manipulation of Host Behavior: An Update and Frequently Asked Questions. in Advances in the Study of Behavior (eds. Brockmann, H. J. et al.) vol. 41 151–186 (Academic Press, 2010).